

EVOLUCIJA GENOME
genetika prirode
Visokokvalitetni sklop genoma naglašava karakteristike genoma raži i agronomski važne gene
PacBio |Illumina |Bionano optička karta |Hi-C Genome Assembly |Genetska karta |Selektivni pregledi |RNA-Seq |ISO-seq |SLAF-seq
Biomarker Technologies je pružio tehničku podršku za Pacbio sekvenciranje, Hi-C sekvenciranje i analizu podataka u ovoj studiji.
Highlights
1. Dobijen je prvi visokokvalitetni genom raži na hromozomskom nivou, koji ima veličinu jednog hromozoma veću od 1 Gb.
2. U poređenju sa Tu, Aet i Hv genomom, jedinstveni nedavni LTR-RT događaji uočeni su u genomu raži, koji je bio odgovoran za proširenje veličine genoma raži.
3. Divergencija između raži i diploidne pšenice dogodila se nakon odvajanja ječma od pšenice, pri čemu su vremena divergencije za dva događaja bila približno 9,6 i 15 MYA.
Fosforilacija FT gena može kontrolisati svojstvo ranog uzgoja u raži.
4. Analiza selektivnog pregleda ukazuje na moguću uključenost ScID1 u regulaciju datuma odrednice i njegov vjerojatni odabir pripitomljavanjem u raži
Pozadina
Pozadina
Raž je vrijedna prehrambena i krmna kultura, važan genetski resurs za poboljšanje pšenice i tritikalea i nezamjenjiv materijal za efikasne komparativne genomičke studije trava.Weining raž, ranocvjetna sorta uzgajana u Kini, izvanredna je zbog svoje otpornosti širokog spektra na pepelnicu i prugastu rđu.Da bismo razumjeli genetsku i molekularnu osnovu elitnih osobina raži i promovirali genomske i oplemenjivačke studije u raži i srodnim usjevima, ovdje smo sekvencirali i analizirali genom Weining raži.
Dostignuća
Genom raži
Genom Raži je konstruisan češljanjem PacBio SMRT očitavanja, kratkog čitanja Illumina sekvenciranja, kao i onih iz hvatanja konformacije hromatina (Hi-C), genetskog mapiranja i BioNano analize.Sakupljeni kontigi (7,74 Gb) činili su 98,47% procijenjene veličine genoma (7,86 Gb), sa 93,67% kontiga (7,25 Gb) dodijeljenih sedam hromozoma.Ponavljajući elementi su činili 90,31% sastavljenog genoma.
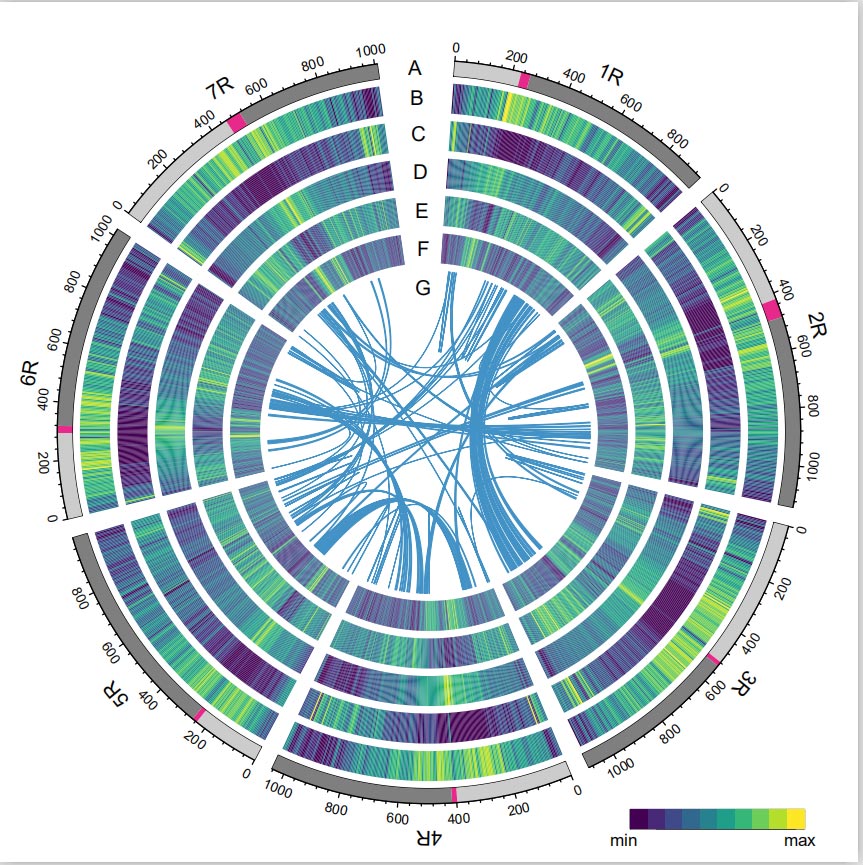
Genom raži
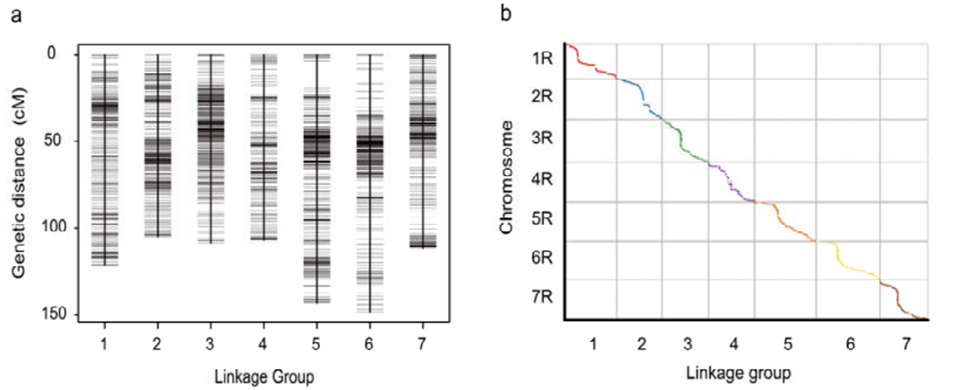
Mapa genetičke veze (WJ) razvijena korištenjem 295 F2 biljaka dobivenih ukrštanjem dviju rasa raži (Weining × Jingzhou)
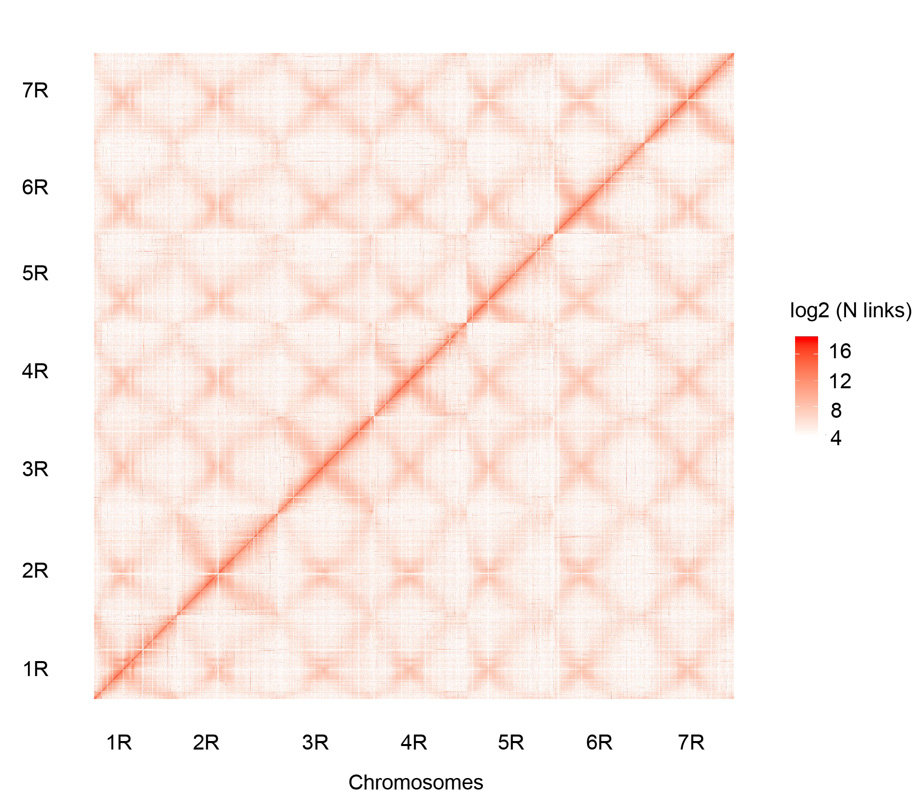
Hi-C kontakt mapa od sedam sastavljenih Weining hromozoma raži (1R – 7R)
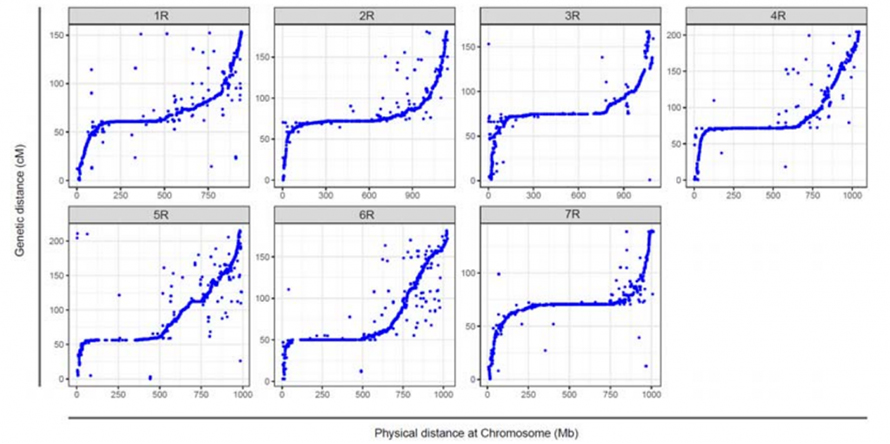
Poravnanje između sedam sklopljenih hromozoma Weining raži i sedam vezanih grupa raži razvijeno korištenjem Lo7 x Lo255 RIL populacije
Utvrđeno je da je LTR Assembly Index (LAI) genoma raži 18,42 i 1393 (96,74%) od 1440 visoko konzerviranih BUSCO gena je identificirano. Ovi rezultati sugeriraju da je sekvenca Weiningovog genoma raži visokog kvaliteta u oba međugena i genske regije.Predviđeno je ukupno 86.991 gena koji kodiraju proteine, uključujući 45.596 gena visoke pouzdanosti (HC) i 41.395 gena niske pouzdanosti (LC).
2. Analiza TE
Analiza TE.Ukupno 6,99 Gb, što predstavlja 90,31% Weining sklopa, označeno je kao TE, što je uključivalo 2,671,941 elementa koji pripadaju 537 porodica.Ovaj sadržaj TE bio je jasno veći od prethodnog prijavljenog za Ta (84,70%), Tu (81,42%), Aet (84,40%), WEW (82,20%) ili Hv (80,80%).Retrotranspozoni sa dugim terminalnim ponavljanjem (LTR-RT), uključujući Gypsy, Copia i neklasifikovane RT elemente, bili su dominantni TE, i 1 su zauzimali 84,49% anotiranog sadržaja TE i 76,29% sastavljenog Weining genoma;CACTA DNK transpozoni bili su drugi najzastupljeniji TE, koji čine 11,68% označenog sadržaja TE i 10,55% sastavljenog Weining genoma.
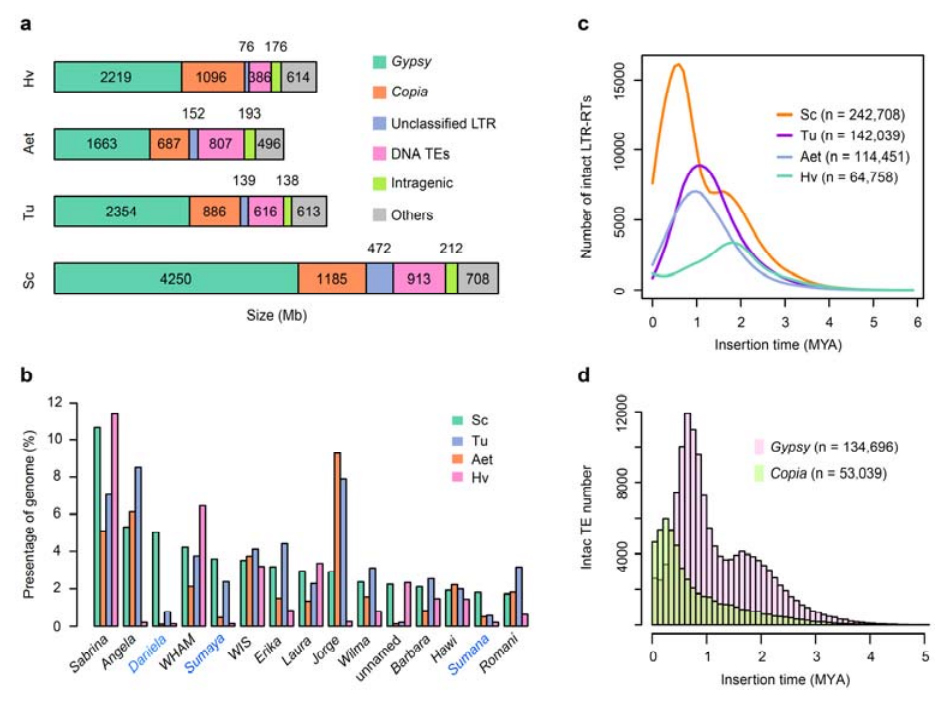
Analiza transpozonskih elemenata raži
Weining raž je imala relativno visok udio nedavnih umetanja LTR-RT-a sa vrhuncem pojačanja koji se pojavio prije oko 0,5 miliona godina (MYA), što je bila najnovija među četiri vrste;drugi vrh, koji se dogodio otprilike 1,7 MYA, bio je stariji i također je viđen kod ječma.Na nivou superfamilije, pronađeni su vrlo nedavni naboji Copia elemenata u Weining raži na 0,3 MYA, dok su pojačanja Gypsy RT-a dominantno oblikovala bimodalni obrazac distribucije LTR-RT dinamike praska.
3. Istraživanje evolucije genoma raži i sintenije hromozoma
Divergencija između raži i diploidne pšenice dogodila se nakon odvajanja ječma od pšenice, pri čemu su vremena divergencije za dva događaja bila približno 9,6 odnosno 15 MYA.1R, 2R, 3R su bili potpuno kolinearni sa grupama 1, 2 i 3 hromozoma pšenice, respektivno.4R, 5R, 6R, 7R su pronađene velike fuzije i segmenti.
4. Analiza duplikacija gena i njihov utjecaj na gene biosinteze skroba
Značajno je da je broj tandemalno dupliciranih gena (TDG) i proksimalno dupliciranih gena (PDG) Weining raži bio veći od onih pronađenih za Tu, Aet, Hv, Bd i Os.Transponirani duplicirani geni (TrDG) su također bili brojniji od onih koji su specifično pronađeni za Tu i Aet.Širenje genoma raži je praćeno većim brojem duplikacija gena.Povećani udari TE u raži su možda doveli do povećanog broja TrDG.
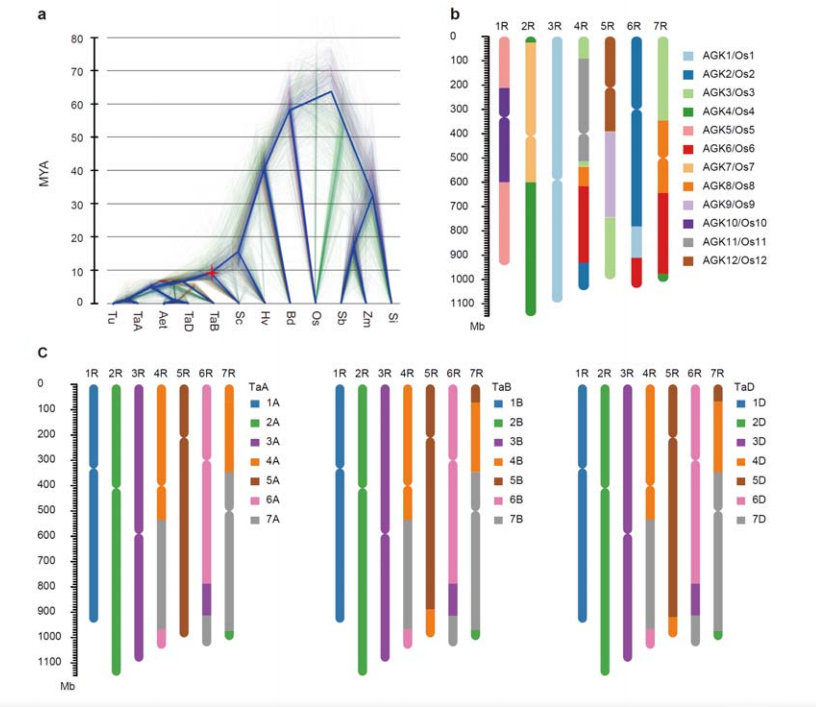
Evolucijska i hromozomska sintezna analiza genoma raži
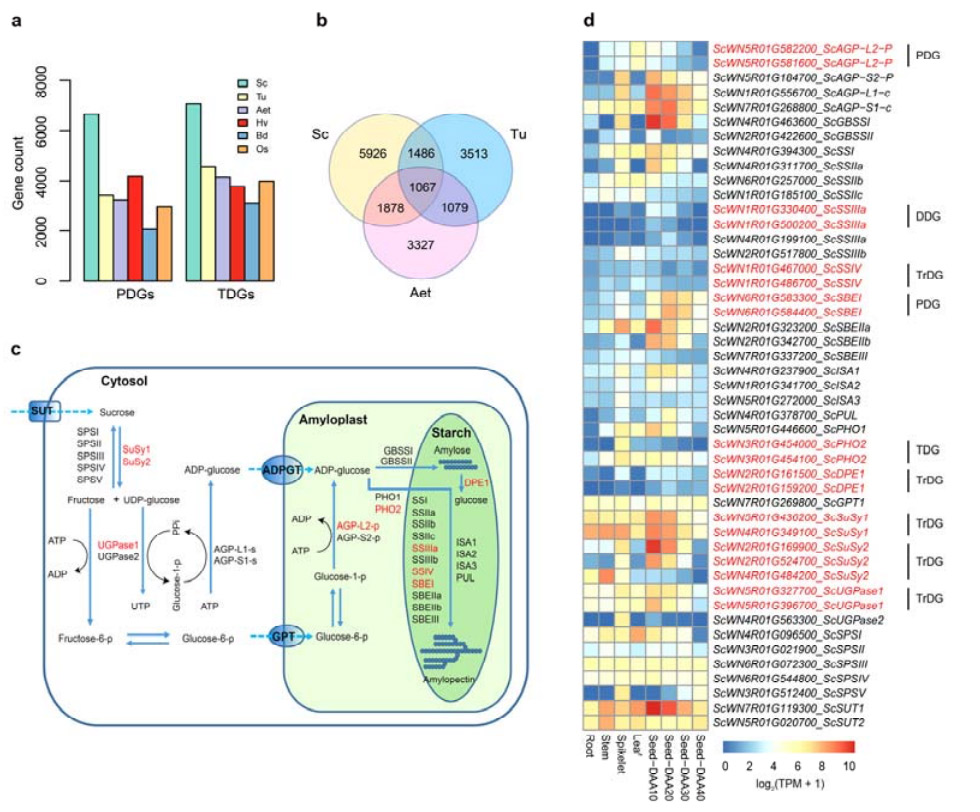
Analiza duplikacija gena raži i njihov utjecaj na raznolikost gena povezanih s biosintezom škroba (SBRG)
5. Disekcija genskih lokusa proteina za skladištenje sjemena raži (SSP).
Četiri hromozomska lokusa (Sec-1 do Sec-4) koji specificiraju raž SSP su identifikovana na 1R ili 2R.α-gliadin geni su evoluirali tek nedavno u pšenici i srodnim vrstama nakon odstupanja pšenice od raži.
6. Ispitivanje transkripcionog faktora (TF) i gena otpornosti na bolesti
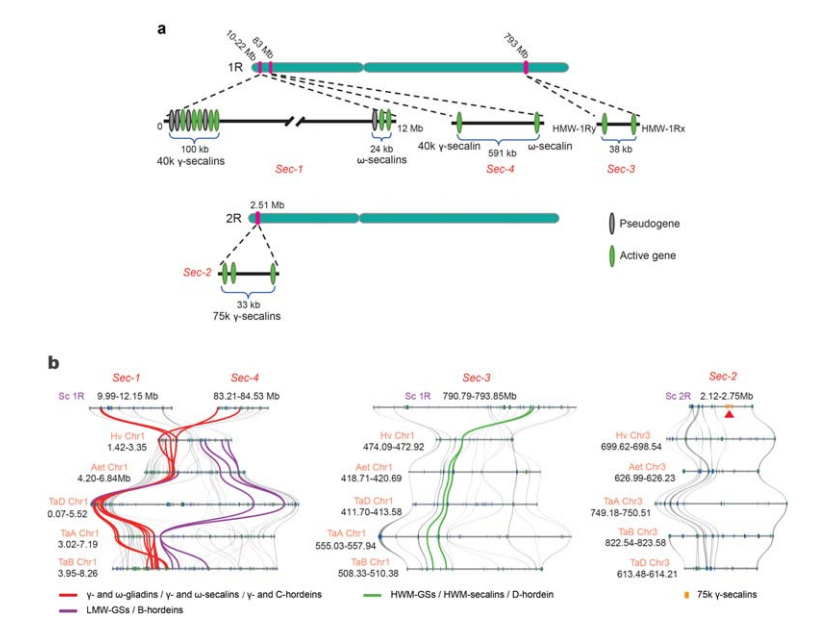
Analiza sekalinskih lokusa raži
Weining raž je imala više gena povezanih s otpornošću na bolesti (DRA) (1.989, dodatni podaci 3) od Tu (1.621), Aet (1.758), Hv (1.508), Bd (1.178), Os (1.575) i A (1.836). ), B (1.728) i D (1.888) podgenomi obične pšenice.
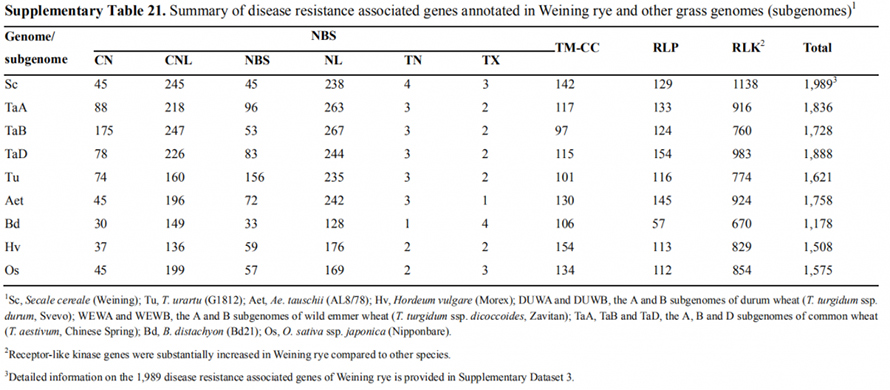
7. Istraživanje osobina ekspresije gena povezanih sa ranim svojstvom glave
Dva FT gena sa relativno visokom ekspresijom u uslovima dugog dana, ScFT1 i ScFT2, označena su u Weiningovom skupu genoma.Dva aminokiselinska ostatka fosforilacije ScFT2 (S76 i T132) pronađena su u vezi sa smanjenjem vremena kontrole
Karakteristike razvoja i ekspresije gena povezane sa ranim svojstvom roda Weining raži
8. Iskopavanje hromozomskih regija i lokusa potencijalno uključenih u pripitomljavanje raži
Ukupno 123.647 SNP-ova korišteno je za provođenje selektivne analize između kultivirane raži i S. vavilovii.11 selektivnih sweep signala identificiranih indeksom redukcije (DRI), indeksom fiksacije (FST) i XP-CLR metodom.Utvrđeno je da je ScID1 moguća umiješanost u regulaciju datuma naslova.
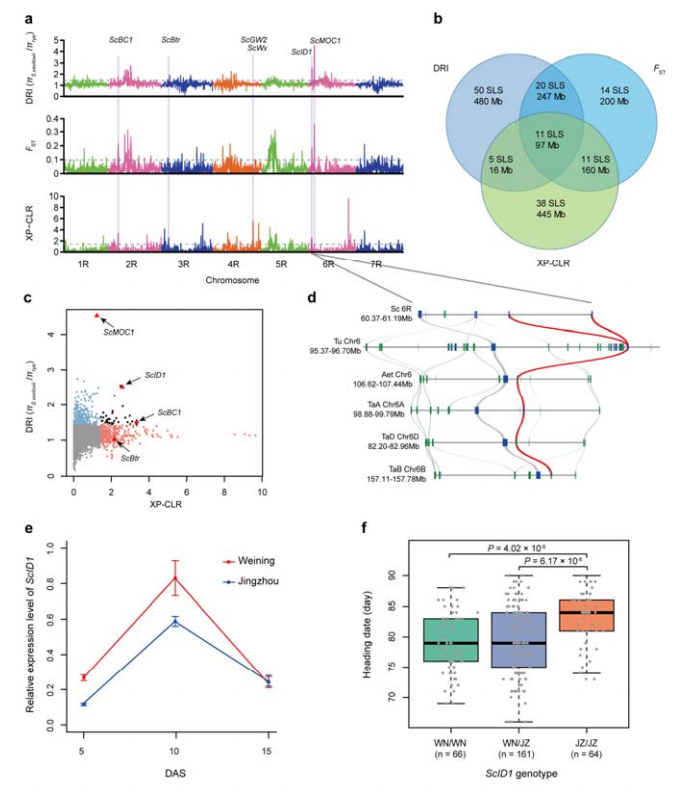
Identifikacija i analiza hromozomskih regija i lokusa potencijalno povezanih s pripitomljavanjem raži
Referenca
Li GW et al.Visokokvalitetni sklop genoma naglašava karakteristike genoma raži i agronomski važne gene.Prirodna genetika (2021)
Vijesti i istaknuti detalji ima za cilj dijeljenje najnovijih uspješnih slučajeva sa Biomarker Technologies, bilježi nova naučna dostignuća, kao i istaknute tehnike primijenjene tokom studije.
Vrijeme objave: Jan-05-2022