

ЕВОЛЮЦІЯ ГЕНОМА
генетика природи
Високоякісна збірка геному висвітлює геномні характеристики жита та агрономічно важливі гени
PacBio |Illumina |Оптична карта Bionano |Збірка геному Hi-C |Генетична карта |Вибіркові розгортки |RNA-Seq |ISO-seq |SLAF-послідовність
Компанія Biomarker Technologies надала технічну підтримку щодо секвенування Pacbio, секвенування Hi-C та аналізу даних у цьому дослідженні.
Основні моменти
1. Було отримано перший високоякісний геном жита на хромосомному рівні, розмір однієї хромосоми якого перевищує 1 Гб.
2. Порівняно з геномами Tu, Aet і Hv, нещодавні унікальні події LTR-RT спостерігалися в геномі жита, що відповідало за збільшення розміру геному жита.
3. Розбіжність між житом і диплоїдними пшеницями відбулася після відділення ячменю від пшениці, причому час розбіжності для двох подій становив приблизно 9,6 і 15 млн років тому.
Фосфорилювання генів FT може контролювати ранню ознаку колосіння жита.
4. Вибірковий аналіз вказує на можливу участь ScID1 у регулюванні дати вирощування та його можливий відбір шляхом одомашнення жита
Фон
Фон
Жито — цінна продовольча та кормова культура, важливий генетичний ресурс для поліпшення пшениці та тритикале, незамінний матеріал для ефективних порівняльних досліджень геноміки трав.Жито Вейнінг, ранньоцвітучий сорт, культивований у Китаї, є видатним завдяки своїй стійкості широкого спектру як до борошнистої роси, так і до смугастої іржі.Щоб зрозуміти генетичну та молекулярну основу елітних ознак жита та сприяти геномним та селекційним дослідженням жита та споріднених культур, ми тут секвенували та проаналізували геном жита Вейнінга.
Досягнення
Геном жита
Геном Rye був створений шляхом комбінування зчитувань PacBio SMRT, секвенування короткого зчитування Illumina, а також з захоплення конформації хроматину (Hi-C), генетичного картування та аналізу BioNano.Зібрані контиги (7,74 Гб) становлять 98,47% від оціненого розміру геному (7,86 Гб), при цьому 93,67% контигів (7,25 Гб) віднесені до семи хромосом.Повторювані елементи становили 90,31% зібраного геному.
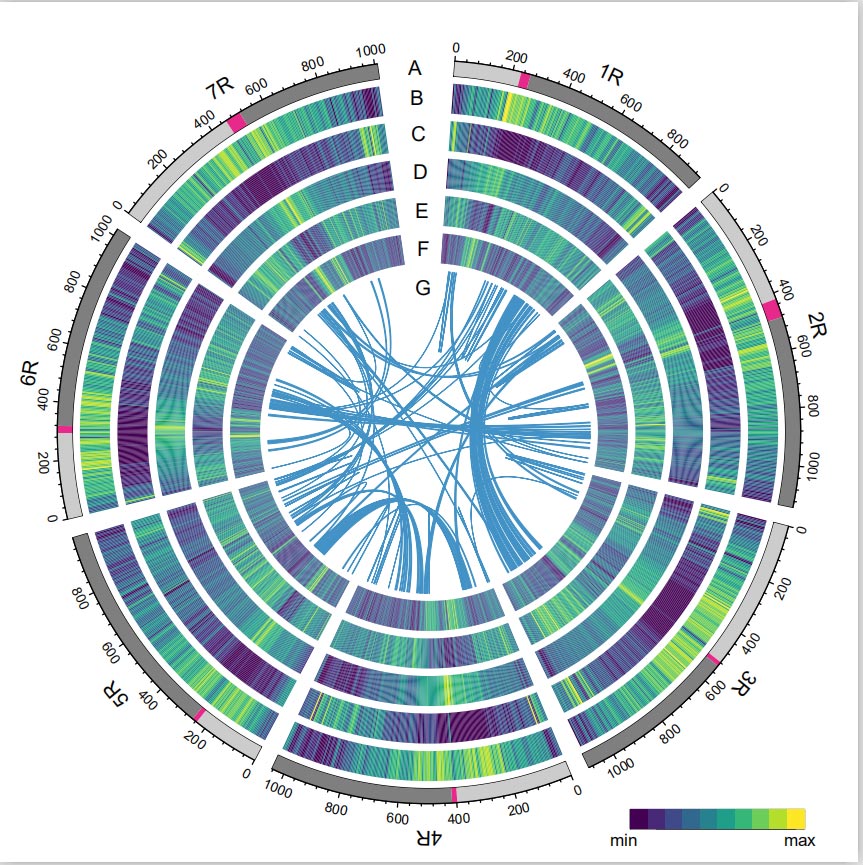
Геном жита
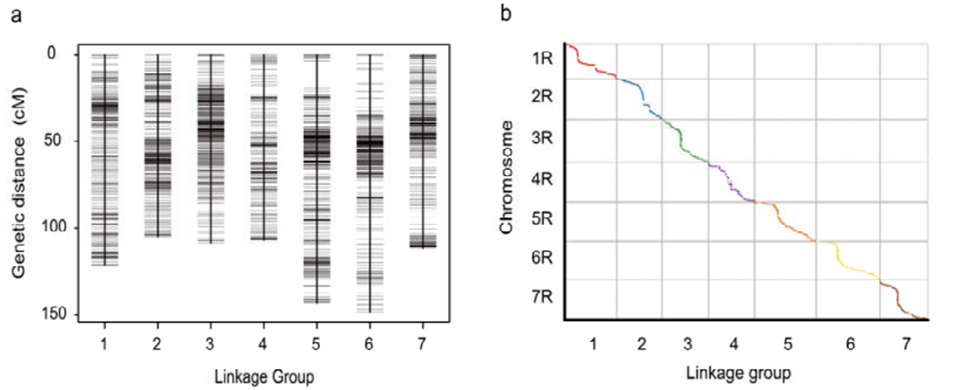
Карта генетичного зчеплення (WJ), розроблена з використанням 295 рослин F2, отриманих від схрещування двох місцевих сортів жита (Weining × Jingzhou)
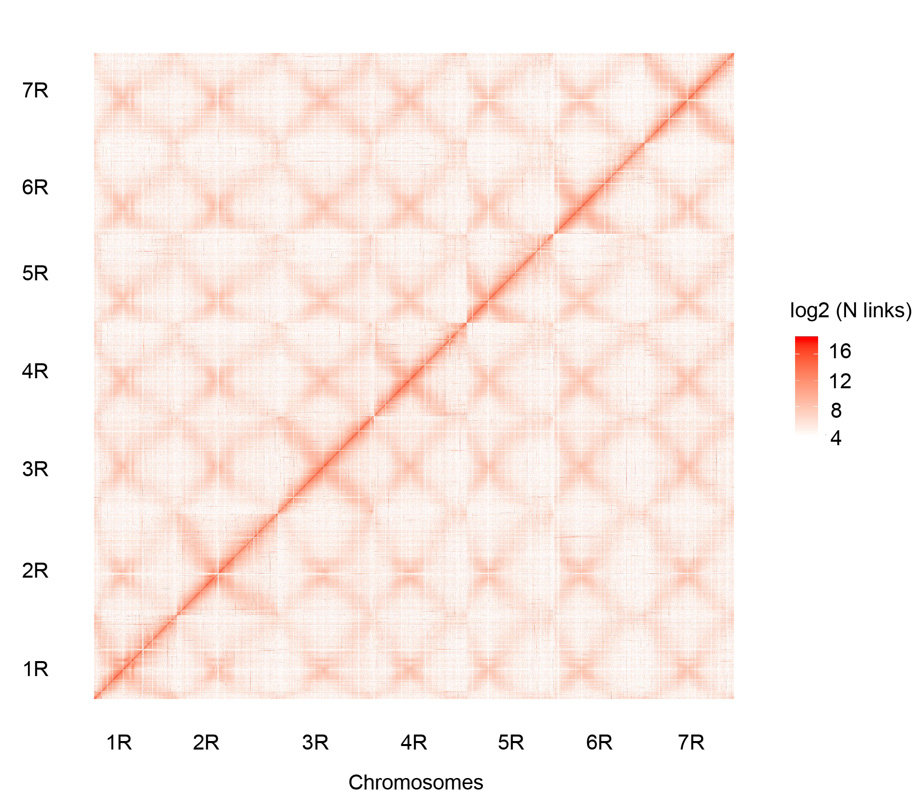
Контактна карта Hi-C семи зібраних хромосом Вейнінга (1R – 7R)
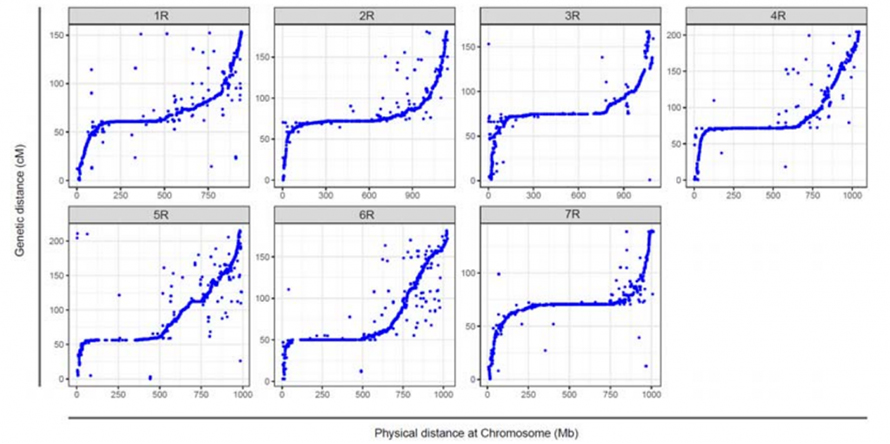
Вирівнювання між сімома зібраними хромосомами жита Вейнінга та сімома групами зчеплення жита, розробленими за допомогою популяції Lo7 x Lo255 RIL
Встановлено, що значення LTR Assembly Index (LAI) геному жита становить 18,42, і 1393 (96,74%) із 1440 висококонсервативних генів BUSCO були ідентифіковані. Ці результати свідчать про те, що послідовність генома жита Вейнінга є високоякісною в обох міжгенних і генні області.Загалом було передбачено 86 991 генів, що кодують білки, включаючи 45 596 генів високої достовірності (HC) і 41 395 генів генів низької достовірності (LC).
2. Аналіз ТЕ
Аналіз ТЕ.Загалом 6,99 Гб, що становить 90,31% складання Weining, було анотовано як TE, що включало 2 671 941 елемент, що належить до 537 сімейств.Цей вміст TE був явно вищим, ніж раніше повідомлялося для Ta (84,70%), Tu (81,42%), Aet (84,40%), WEW (82,20%) або Hv (80,80%).Ретротранспозони з довгим термінальним повторенням (LTR-RT), включаючи Gypsy, Copia та некласифіковані елементи RT, були домінуючими TE, і 1 займав 84,49% анотованого вмісту TE та 76,29% зібраного геному Вейнінга;Транспозони CACTA ДНК були другим за поширеністю TE, складаючи 11,68% анотованого вмісту TE та 10,55% зібраного геному Вейнінга.
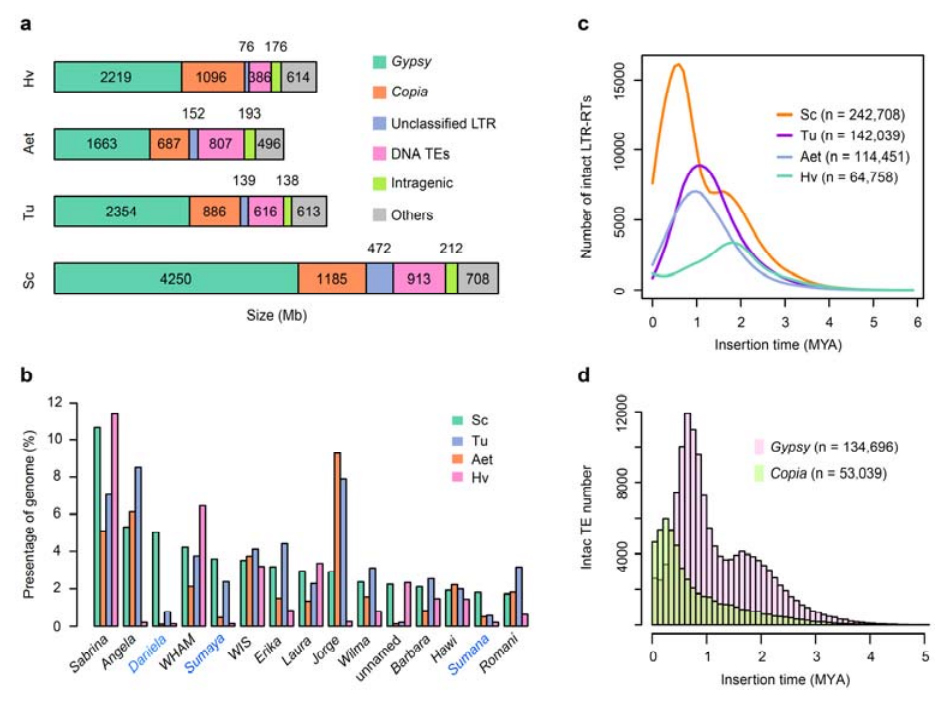
Аналіз транспозонних елементів жита
Жито проростання мало порівняно високу частку останніх інсерцій LTR-RTs з піком ампліфікації, який з’явився приблизно 0,5 мільйона років тому (MYA), що було найновішим серед чотирьох видів;інший пік, який стався приблизно через 1,7 млн років тому, був старшим і також спостерігався в ячмені.На рівні суперродини було виявлено зовсім нещодавні спалахи елементів Copia у житі Weining на 0,3 MYA, тоді як ампліфікації Gypsy RT домінантно сформували бімодальну структуру розподілу динаміки спалахів LTR-RT.
3. Дослідження еволюції геному жита та синтенії хромосом
Розбіжність між житом і диплоїдними пшеницями відбулася після відділення ячменю від пшениці, причому час розбіжності для двох подій становив приблизно 9,6 і 15 млн років тому відповідно.1R, 2R, 3R були повністю колінеарними з групами 1, 2 і 3 хромосом пшениці відповідно.4R, 5R, 6R, 7R виявлено існування великомасштабних злиттів і сегментів.
4. Аналіз подвоєння генів та їх вплив на гени біосинтезу крохмалю
Примітно, що кількість тандемно дубльованих генів (TDG) і проксимально дубльованих генів (PDG) жита Weining була вищою, ніж знайдена для Tu, Aet, Hv, Bd і Os.Транспоновані дубльовані гени (TrDG) також були більш численними, ніж ті, що були спеціально знайдені для Tu та Aet.Розширення геному жита супроводжується більшою кількістю дублікацій генів.Збільшення спалахів TE в житі могло призвести до підвищеної кількості TrDG.
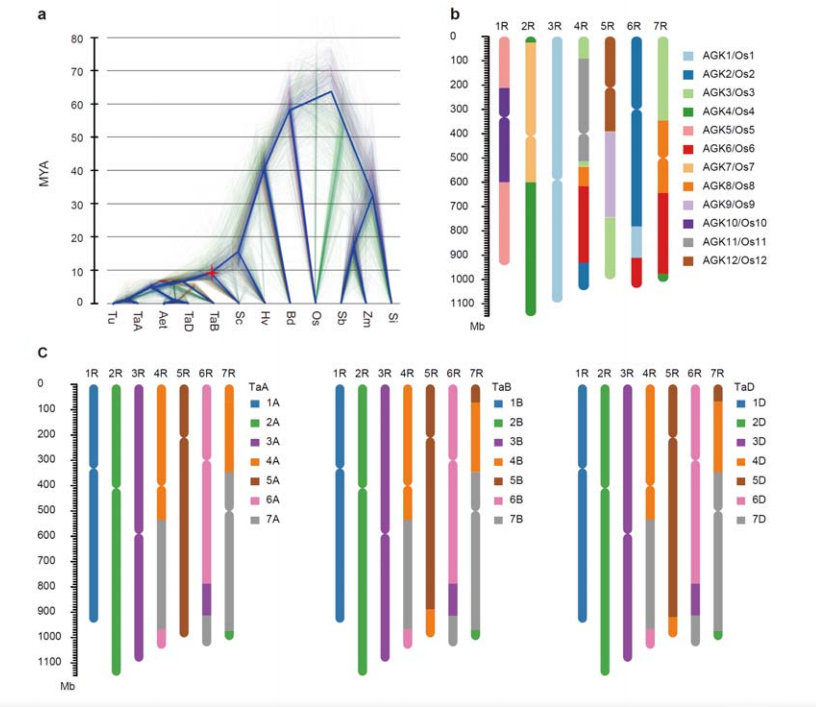
Еволюційний та хромосомно-синтетичний аналіз геному жита
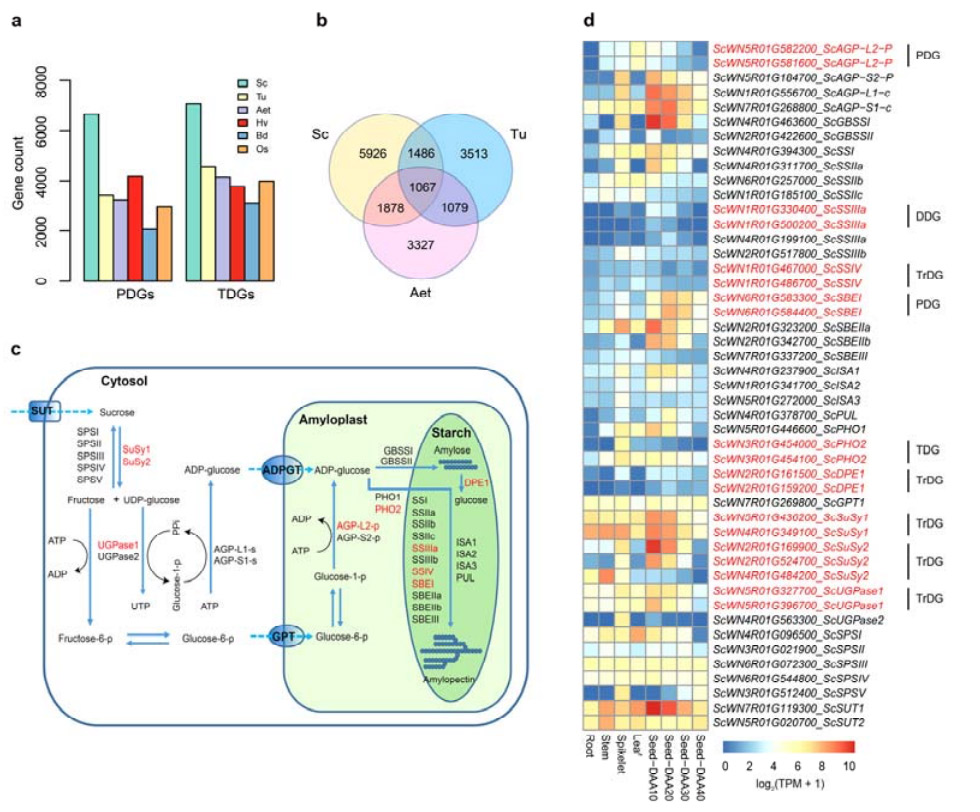
Аналіз подвоєння генів жита та їх вплив на різноманітність генів, пов’язаних з біосинтезом крохмалю (SBRG)
5. Розтин локусів гена запасного білка (SSP) насіння жита
Чотири хромосомні локуси (від Sec-1 до Sec-4), що визначають SSP жита, були ідентифіковані на 1R або 2R.Гени α-гліадину виникли лише нещодавно у пшениці та близькоспоріднених видів після розходження пшениці з жита.
6. Дослідження фактора транскрипції (ТФ) і генів стійкості до хвороб
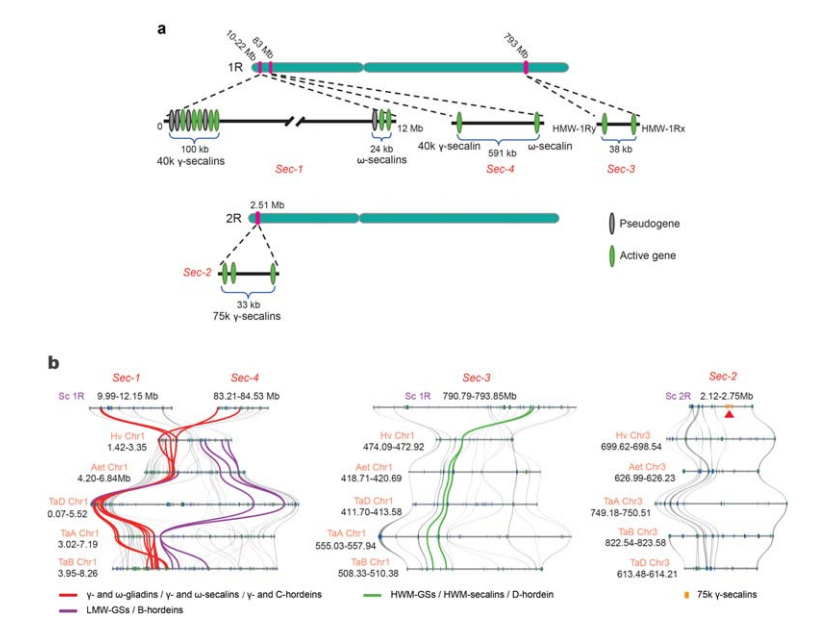
Аналіз локусів секаліну жита
Жито в’янення мало більше генів стійкості до хвороб (DRA) (1989, Додаткові дані 3), ніж Tu (1621), Aet (1758), Hv (1508), Bd (1178), Os (1575) і A (1836). ), B (1728) і D (1888) субгеномів м’якої пшениці.
7. Дослідження особливостей експресії генів, пов'язаних з ранньою ознакою головчатості
Два гени FT з відносно високою експресією в умовах тривалого дня, ScFT1 і ScFT2, були анотовані в збірці геному Вейнінга.Було виявлено взаємозв’язок двох амінокислотних залишків фосфорилювання ScFT2 (S76 і T132) із зниженням контролю часу
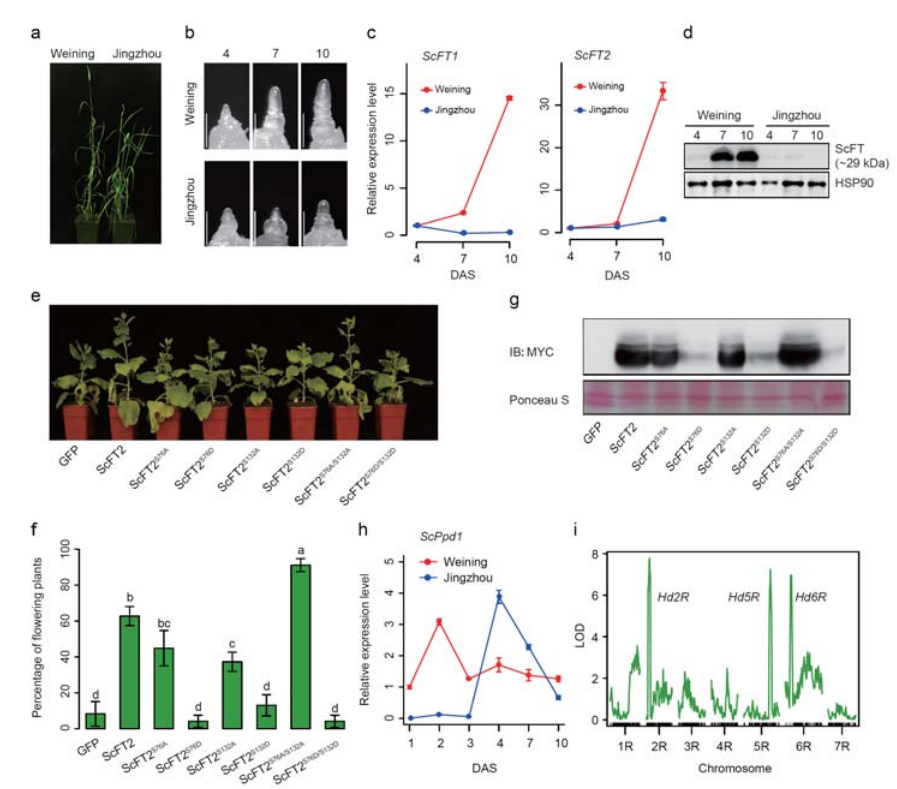
Особливості розвитку та експресії генів, пов’язані з ознакою раннього колошання жита Вейнінга
8. Виявлення хромосомних ділянок і локусів, потенційно залучених до одомашнення жита
Загалом 123 647 SNP було використано для проведення селективного аналізу культивованого жита та S. vavilovii.11 селективних сигналів розгортки, ідентифікованих індексом зменшення (DRI), індексом фіксації (FST) і методом XP-CLR.Було виявлено можливу участь ScID1 у регулюванні дати заголовка.
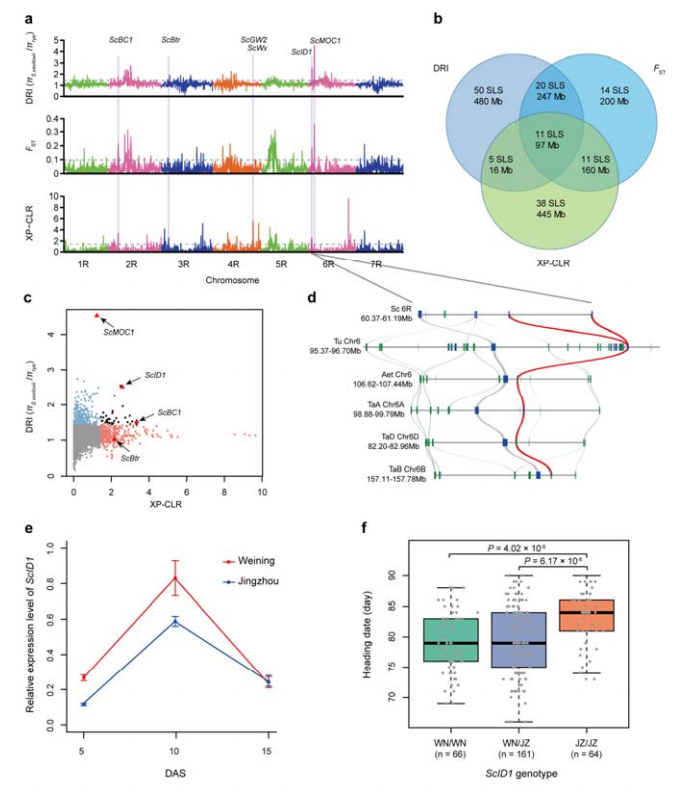
Ідентифікація та аналіз хромосомних областей і локусів, потенційно пов'язаних з одомашненням жита
довідка
Li GW та ін.Високоякісна збірка геному висвітлює геномні характеристики жита та агрономічно важливі гени.Генетика природи (2021)
Новини та основні моменти спрямований на обмін останніми успішними випадками з Biomarker Technologies, фіксацію нових наукових досягнень, а також видатних методів, застосованих під час дослідження.
Час публікації: 05 січня 2022 р