

ЕВОЛУЦИЈА ГЕНОМЕ
генетика природе
Висококвалитетни склоп генома истиче карактеристике генома ражи и агрономски важне гене
ПацБио |Иллумина |Бионано Оптицал Мап |Хи-Ц Геноме Ассембли |Генетска карта |Селецтиве Свеепс |РНА-Сек |ИСО-сек |СЛАФ-сек
Биомаркер Тецхнологиес је пружио техничку подршку за Пацбио секвенцирање, Хи-Ц секвенцирање и анализу података у овој студији.
Хигхлигхтс
1. Добијен је први висококвалитетни геном ражи на хромозомском нивоу, који има величину једног хромозома већу од 1 Гб.
2. У поређењу са Ту, Ает и Хв геномом, јединствени недавни ЛТР-РТ догађаји су примећени у геному ражи, који је био одговоран за проширење величине генома ражи.
3. Дивергенција између ражи и диплоидне пшенице настала је након одвајања јечма од пшенице, при чему су времена дивергенције за два догађаја била приближно 9,6 и 15 МИА.
Фосфорилација ФТ гена може контролисати рану особину у ражи.
4. Анализа селективног прегледа указује на могућу умешаност СцИД1 у регулисање датума одреднице и његову вероватну селекцију припитомљавањем у ражи
Позадина
Позадина
Раж је вредан прехрамбени и крмни усев, важан генетски ресурс за побољшање пшенице и тритикалеа и незаменљив материјал за ефикасна упоредна истраживања геномике трава.Веининг раж, раноцветна сорта која се узгаја у Кини, изванредна је због своје отпорности широког спектра на пепелницу и пругасту рђу.Да бисмо разумели генетску и молекуларну основу елитних особина ражи и промовисали геномске и оплемењивачке студије у ражи и сродним културама, овде смо секвенцирали и анализирали геном Веининг ражи.
Достигнућа
Геном ражи
Геном ражи је конструисан чешљањем ПацБио СМРТ очитавања, кратког читања Иллумина секвенцирања, као и оних из хватања конформације хроматина (Хи-Ц), генетског мапирања и БиоНано анализе.Сакупљени контиги (7,74 Гб) чинили су 98,47% процењене величине генома (7,86 Гб), са 93,67% контига (7,25 Гб) додељених седам хромозома.Понављајући елементи су чинили 90,31% састављеног генома.
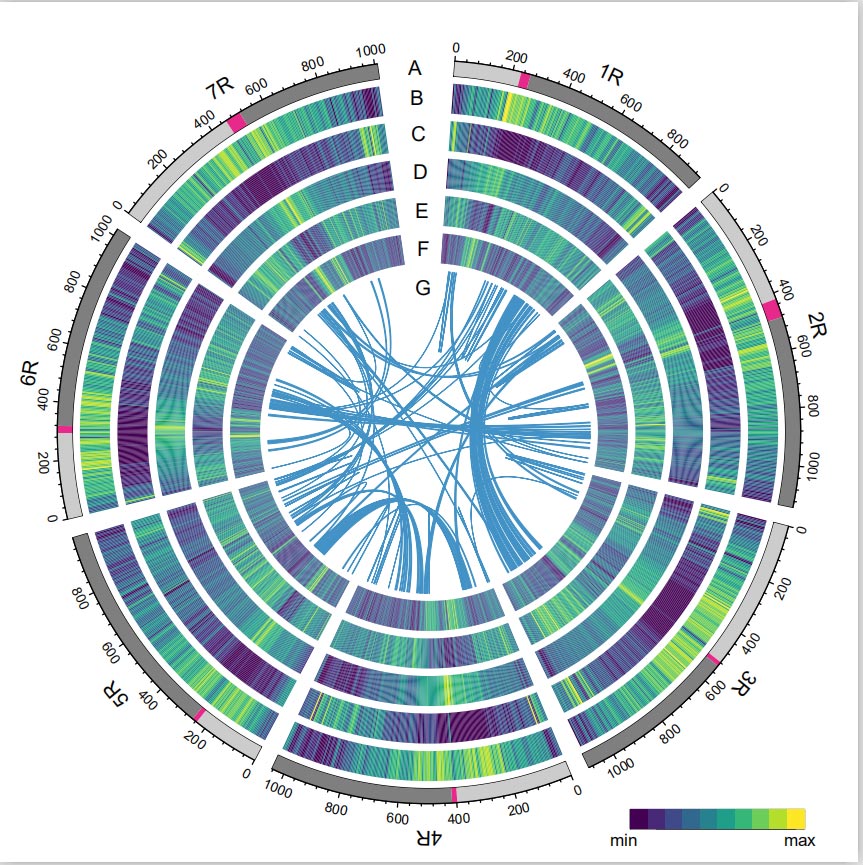
Геном ражи
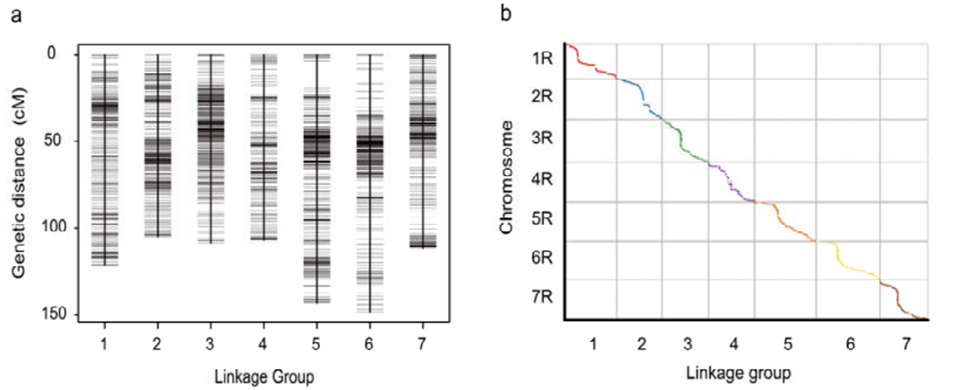
Мапа генетичке везе (ВЈ) развијена коришћењем 295 Ф2 биљака добијених укрштањем две сорте ражи (Веининг × Јингзхоу)
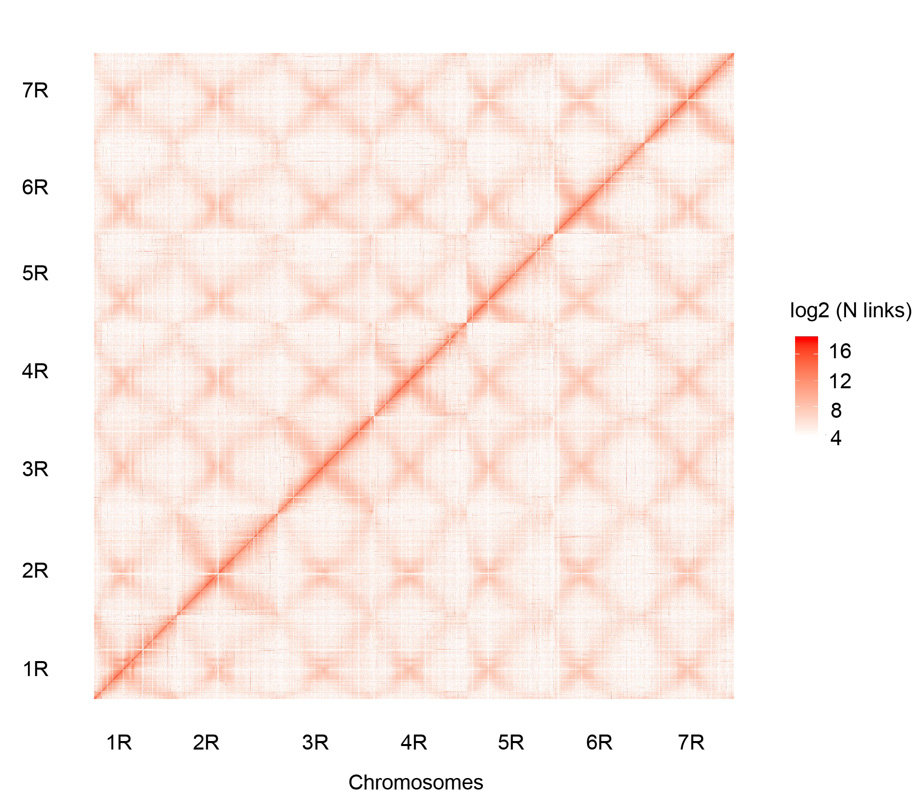
Хи-Ц контакт мапа седам састављених хромозома ражи Веининг (1Р – 7Р)
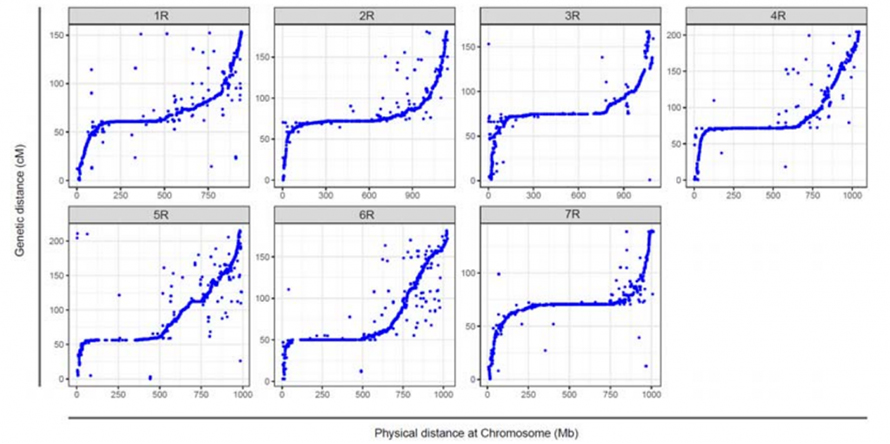
Поравнање између седам склопљених хромозома Веининг ражи и седам везаних група ражи развијено коришћењем Ло7 к Ло255 РИЛ популације
Утврђено је да је ЛТР Ассембли Индек (ЛАИ) генома ражи 18,42 и идентификовано је 1,393 (96,74%) од 1,440 високо конзервираних БУСЦО гена. Ови резултати сугеришу да је секвенца Веининг генома ражи високог квалитета у оба међугена. и генске регије.Предвиђено је укупно 86.991 гена који кодирају протеине, укључујући 45.596 гена високе поузданости (ХЦ) и 41.395 гена са ниском поузданошћу (ЛЦ).
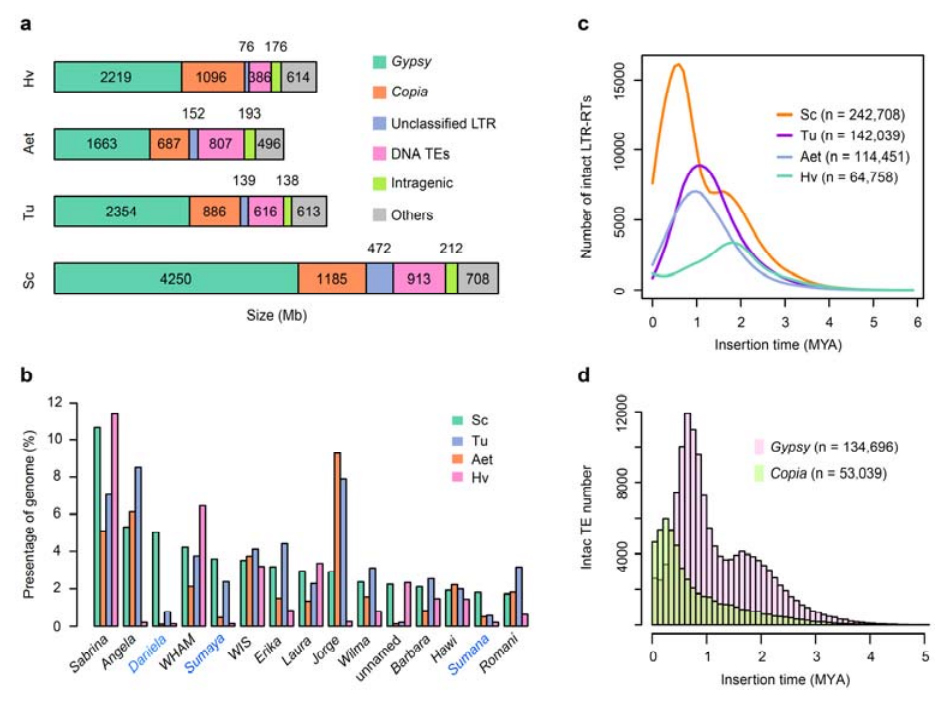
2. Анализа ТЕ
Анализа ТЕ.Укупно 6,99 Гб, што представља 90,31% Веининг склопа, означено је као ТЕ, што је укључивало 2,671,941 елемента који припадају 537 породица.Овај садржај ТЕ био је јасно већи од претходног пријављеног за Та (84,70%), Ту (81,42%), Ает (84,40%), ВЕВ (82,20%) или Хв (80,80%).Ретротранспозони са дугим терминалним понављањем (ЛТР-РТ), укључујући Гипси, Цопиа и некласификоване РТ елементе, били су доминантни ТЕ, и 1 су заузимали 84,49% обележеног садржаја ТЕ и 76,29% састављеног Веининг генома;ЦАЦТА ДНК транспозони су били други најзаступљенији ТЕ, чинећи 11,68% обележеног садржаја ТЕ и 10,55% састављеног Веининг генома.
Анализа транспозонских елемената ражи
Веининг раж је имала релативно висок удео недавних уметања ЛТР-РТ-а са врхунцем појачања који се појавио пре око 0,5 милиона година (МИА), што је била најновија међу четири врсте;други врхунац, који се догодио отприлике 1,7 МИА, био је старији и такође је виђен код јечма.На нивоу суперфамилије, пронађени су врло недавни избијање Цопиа елемената у Веининг ражи на 0,3 МИА, док су појачања Гипси РТ-а доминантно обликовала бимодални образац дистрибуције ЛТР-РТ динамике праска.
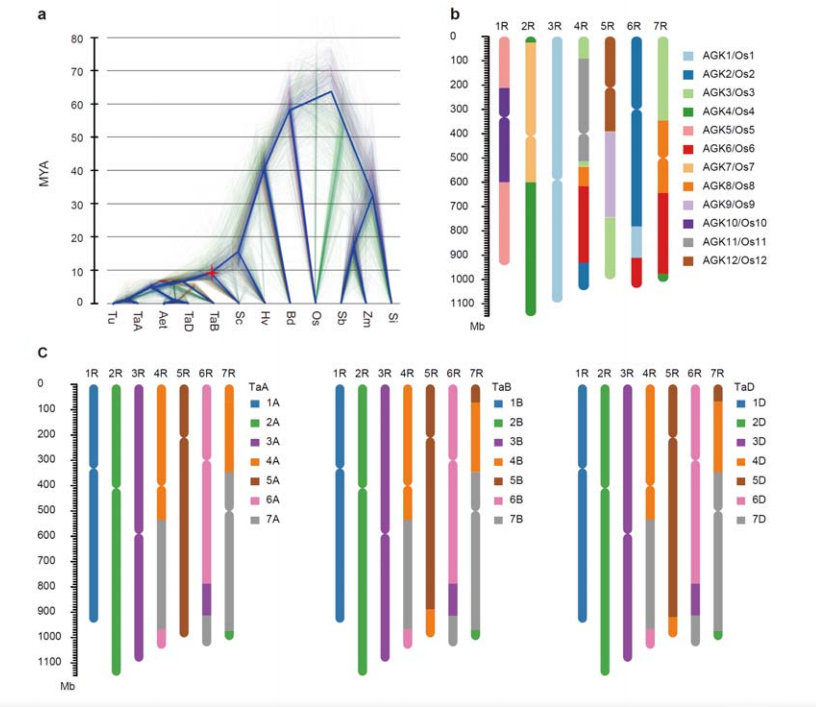
3. Истраживање еволуције генома ражи и синтеније хромозома
Дивергенција између ражи и диплоидне пшенице десила се након одвајања јечма од пшенице, при чему су времена дивергенције за два догађаја била приближно 9,6 односно 15 МИА, респективно.1Р, 2Р, 3Р су били потпуно колинеарни са групама 1, 2 и 3 хромозома пшенице, респективно.Утврђено је да 4Р, 5Р, 6Р, 7Р постоје велике фузије и сегменти.
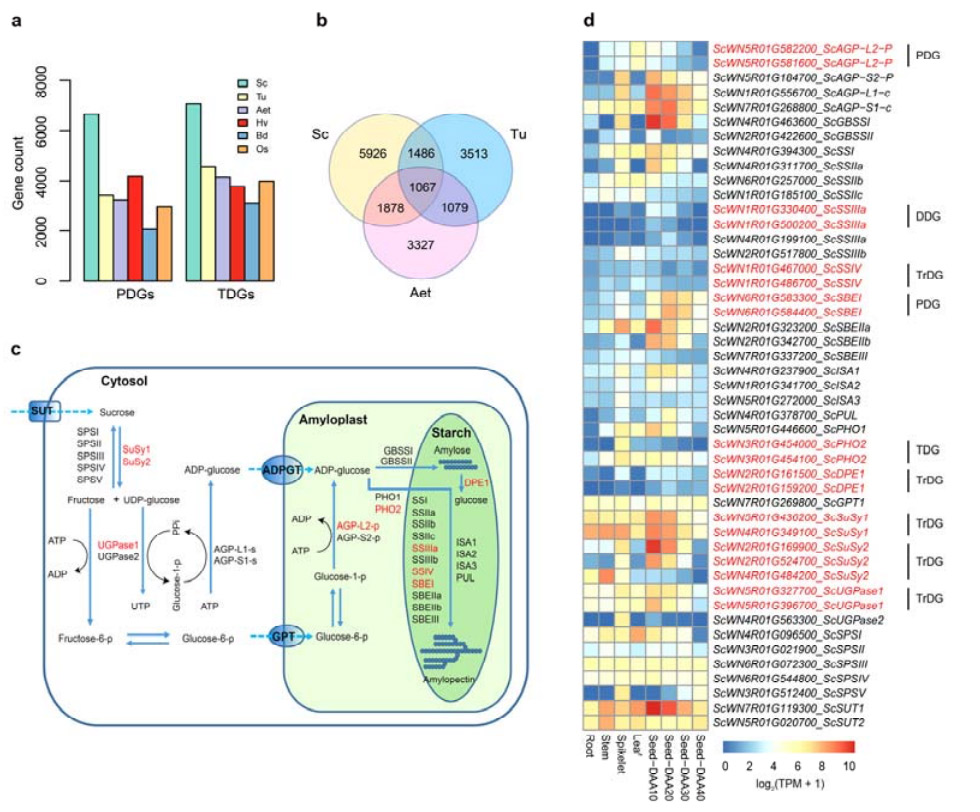
4. Анализа дупликација гена и њихов утицај на гене биосинтезе скроба
Значајно је да је број тандемално дуплицираних гена (ТДГ) и проксимално дуплицираних гена (ПДГ) Веининг ражи био већи од оних пронађених за Ту, Ает, Хв, Бд и Ос.Транспоновани дуплирани гени (ТрДГ) су такође били бројнији од оних који су посебно пронађени за Ту и Ает.Ширење генома ражи је праћено већим бројем дупликација гена.Повећани удари ТЕ у ражи су можда довели до повећаног броја ТрДГ.
Еволуциона и хромозомска синтенија анализа генома ражи
Анализа дупликација гена ражи и њихов утицај на разноликост гена повезаних са биосинтезом скроба (СБРГ)
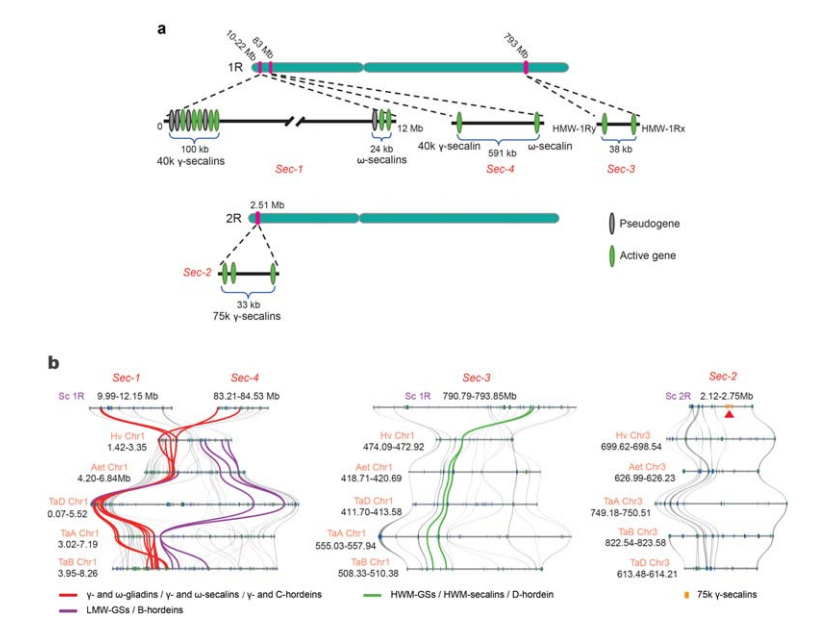
5. Дисекција локуса гена протеина за складиштење семена ражи (ССП).
Четири хромозомска локуса (Сец-1 до Сец-4) који специфицирају раж ССП су идентификована на 1Р или 2Р.α-глиадин гени су еволуирали тек недавно у пшеници и блиско сродним врстама након дивергенције пшенице од ражи.
6. Испитивање транскрипционог фактора (ТФ) и гена отпорности на болести
Анализа секалинских локуса ражи
Веининг раж је имала више гена повезаних са отпорношћу на болести (ДРА) (1.989, додатни подаци 3) од Ту (1.621), Ает (1.758), Хв (1.508), Бд (1.178), Ос (1.575) и А (1.836). ), Б (1.728) и Д (1.888) субгеноми обичне пшенице.
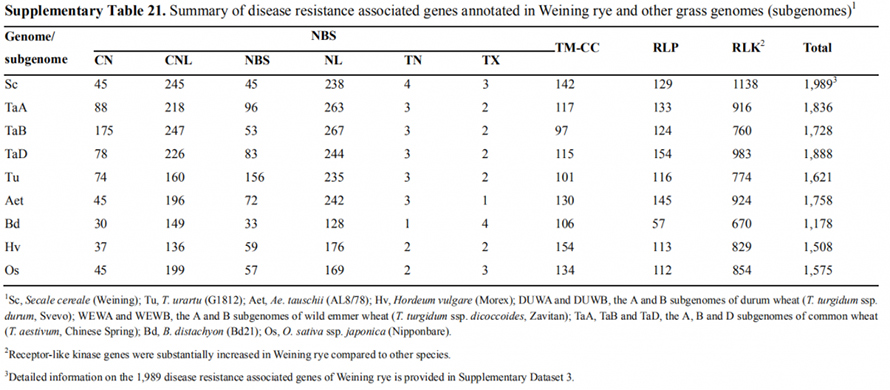
7. Истраживање особина експресије гена повезаних са раним особинама главе
Два ФТ гена са релативно високом експресијом у условима дугог дана, СцФТ1 и СцФТ2 , су означена у Веининговом скупу генома.Два аминокиселинска остатка фосфорилације СцФТ2 (С76 и Т132) пронађена су у вези са контролом времена смањења
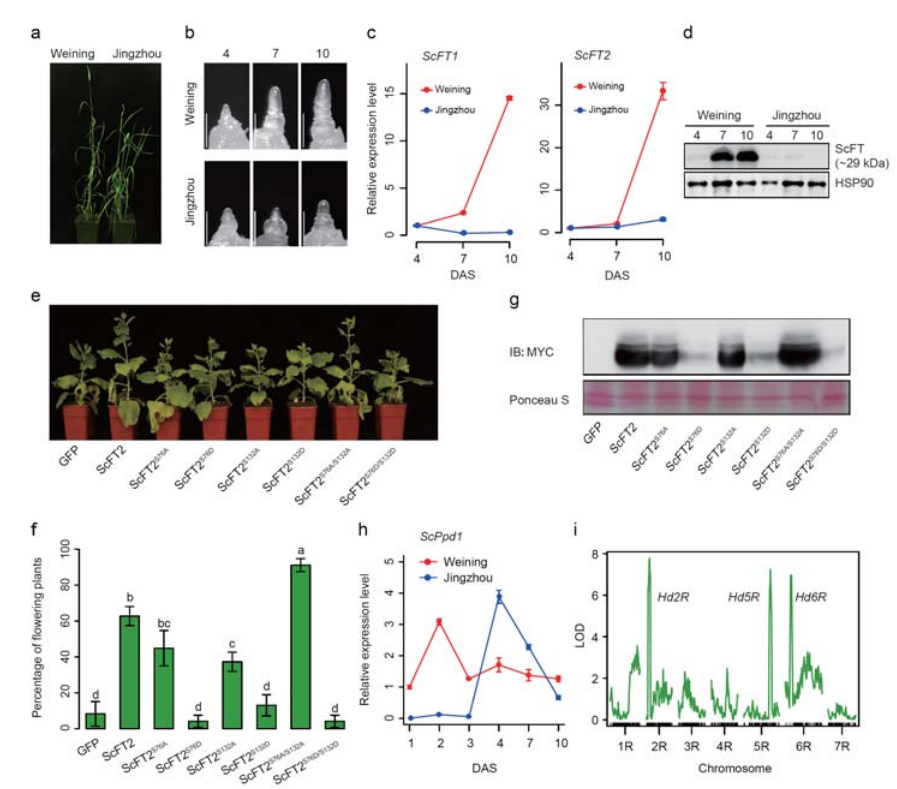
Карактеристике развоја и експресије гена повезане са особином раног грла Веининг ражи
8. Ископавање хромозомских региона и локуса потенцијално укључених у припитомљавање ражи
Укупно 123.647 СНП-а је коришћено за спровођење селективне анализе између култивисане ражи и С. вавиловии.11 селективних свееп сигнала идентификованих индексом редукције (ДРИ), индексом фиксације (ФСТ) и КСП-ЦЛР методом.Утврђено је да је СцИД1 могућа умешаност у регулисање датума наслова.
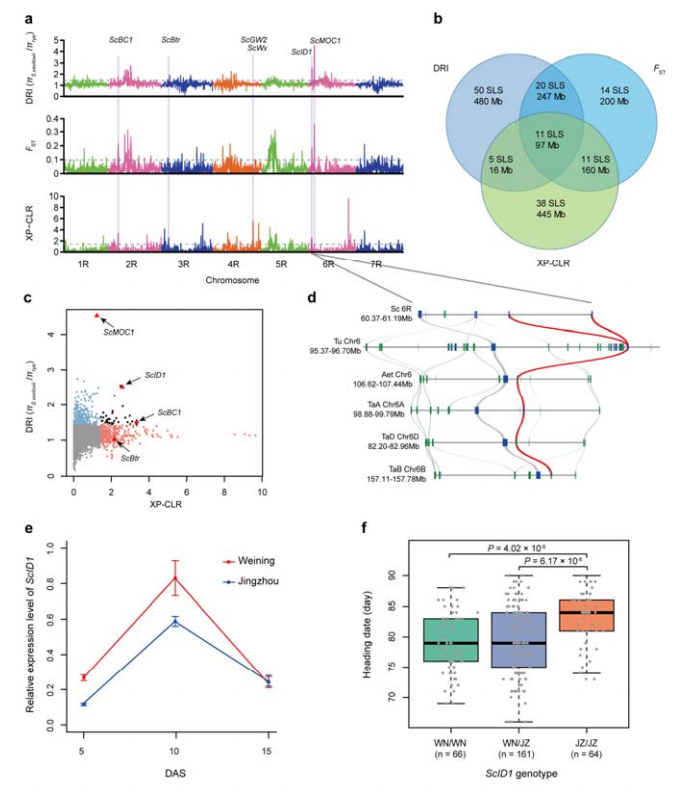
Идентификација и анализа хромозомских региона и локуса потенцијално повезаних са припитомљавањем ражи
Референца
Ли ГВ ет ал.Висококвалитетни склоп генома истиче карактеристике генома ражи и агрономски важне гене.Природна генетика (2021)
Вести и најважније ствари има за циљ да подели најновије успешне случајеве са Биомаркер Тецхнологиес, ухвати нова научна достигнућа, као и истакнуте технике примењене током студије.
Време поста: Јан-05-2022